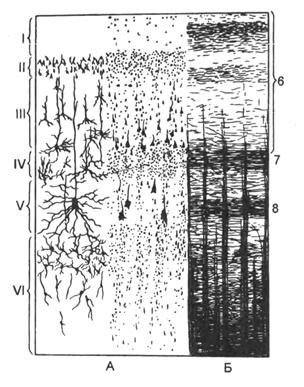
Порядок розташування шарів кори великих півкуль. Кора великих півкуль це
Кора великого мозкупокриває поверхню півкуль і утворює велику кількість різних за глибиною та протяжністю борозен (лат. sulci cerebri). Між борознами розташовані різної величини звивини великого мозку (лат. gyri cerebri) .
У кожній півкулі розрізняють такі поверхні:
Ці три поверхні кожної півкулі, переходячи одна в іншу, утворюють три краї. верхній край (лат. margo superior) розділяє верхньолатеральну та медіальну поверхні. Нижньолатеральний край (лат. margo inferolateralis) відокремлює верхньолатеральну поверхню від нижньої. Нижньомідіальний край (лат. margo inferomedialis) розташовується між нижньою та медіальною поверхнями .
У кожній півкулі розрізняють місця, що найбільш виступають: спереду - лобовий полюс (лат. polus frontalis), ззаду - потиличний (лат. polus occipitalis), і збоку - скроневий (лат. polus temporalis) .
Півкуля розділена на п'ять часток. Чотири з них примикають до відповідних кісток склепіння черепа:
Борозни та звивини верхньолатеральної поверхні
Лобна частка – позначена синім. тім'яна частка - позначена жовтим. Скронева частка - позначена зеленим. Потилична частка- Позначена рожевим.
Лобна частка
Лобну частку від тім'яної відокремлює глибока центральна борозна (лат. sulcus centralis). Вона починається на медіальної поверхніпівкулі, переходить на його верхньолатеральну поверхню, йде по ній трохи косо, ззаду наперед, і зазвичай не доходить до неї латеральної борознимозку.
Приблизно паралельно центральній борозні розташовується передцентральна борозна (лат. sulcus precentralis), яка не доходить до верхнього краю півкулі. Передцентральна борозна облямовує спереду прецентральну звивину (лат. gyrus precentralis) .
Верхня та нижня лобні борозни(Лат. sulci frontales superior et inferior ) прямують від попередньо центральної борознивперед. Вони ділять лобну частку на:
Від латеральної борозни вгору відходять дрібні борозни, звані гілками. Найбільш постійними є висхідна (лат. ramus ascendens) та передня (лат. ramus anterior) Гілки. Верхньозадній відділ борозни називається задньою гілкою (лат. ramus posterior) .
Нижня лобна звивина, в межах якої проходять висхідна та передня гілки, поділяється ними на три частини:
тім'яна частка
Залягає позаду центральної борозни, яка відокремлює її від лобової. Від скроневої відмежована латеральною борозеною мозку, від потиличної - частиною тім'яно-потиличної борозни (лат. sulcus parietooccipitalis) .
Між висхідною і задньою гілками латеральної борозни мозку розташована ділянка кори, що позначається як лобово-тім'яна покришка (лат. operculum frontoparietalis). До неї входять задня частина нижньої лобової звивини, нижні відділи передцентральної та постцентральної звивин, а також нижній відділ передньої частини тім'яної частки.
Скронева частка
Має найбільш виражені межі. У ній розрізняють опуклу латеральну поверхню та увігнуту нижню. Тупий полюс скроневої частки звернений вперед і вниз. Латеральна борозна великого мозку різко відмежовує скроневу частку від лобової.
Дві борозни, розташовані на верхньолатеральної поверхні: верхня (лат. sulcus temporalis superior) та нижня (лат. sulcus temporalis inferior) скроневі борозни, слідуючи майже паралельно латеральної борозні мозку, поділяють частку на три скроневі звивини: верхню, середню та нижню (лат. gyri temporales superior, medius et inferior ) .
Ті ділянки скроневої частки, які спрямовані у бік латеральної борозни мозку порізані короткими поперечними скроневими борознами(Лат. sulci temporales transversi). Між цими борознами залягають 2-3 короткі поперечні скроневі звивини, пов'язані з звивинами скроневої частки (лат. gyri temporales transversi) та острівцем .
Острівцева частка (острівець)
Залягає на дні латеральної ямки величезного мозку (лат. fossa lateralis cerebri).
Вона є тристоронньою пірамідою, зверненою своєю вершиною - полюсом острівця - кпереду і назовні, у бік латеральної борозни. З периферії острівець оточений лобовою, тім'яною та скроневою частинами, що беруть участь в утворенні стінок латеральної борозни мозку.
Основа острівця з трьох сторін оточена круговою борозеною острівця (лат. sulcus circularis insulae).
Його поверхня прорізана глибокої центральної борозна острівця (лат. sulcus centralis insulae). Ця борозна розділяє острівець на передню та задню частини.
На поверхні розрізняють велику кількість дрібних звивин острівця (лат. gyri insulae). Велика передня частина складається з декількох коротких звивин острівця (лат. gyri breves insulae), задня - однією довгою звивиною (лат. gyrus longus insulae) .
Борозни та звивини медіальної поверхні
На медіальну поверхню півкулі виходять лобова, тім'яна та потилична частки.
Поясна звивина обмежена зверху поясною борозна (лат. sulcus cinguli). В останній розрізняють опуклу до лобового полюса передню частину і задню частину, Яка, слідуючи вздовж поясної звивини і не доходячи до її заднього відділу, піднімається до верхнього краю півкулі великого мозку. Задній кінець борозни лежить позаду верхнього кінця центральної борозни. Між передцентральної борозна, закінчення якої іноді добре видно у верхнього краю медіальної поверхні півкулі, і кінцем поясної борозни, розташовується парацентральна часточка (лат. lobulus paracentralis) .
Вище поясної звивини, Спереду від підмозолистого поля, починається медіальна звивина лоба (лат. gyrus frontalis medialis). Вона тягнеться до парацентральної часточки і є нижньою частиноюверхньої лобової звивини.
Ззаду від поясної борозни лежить невелика чотирикутна часточка - передклин (лат. precuneus). Її заднім кордономє глибока тім'яно-потилична борозна (лат. sulcus parietooccipitalis), нижній - підтеменна борозна (лат. sulcus subparietalis), що відокремлює передклиння від заднього відділу поясної звивини.
Ззаду і нижче проклинання залягає трикутна часточка - клин (лат. cuneus). Випукла зовнішня поверхня клину бере участь у освіті потиличного полюса. Спрямована вниз і вперед вершина клину майже доходить до заднього відділу поясної звивини. Заднижньою межею клина є дуже глибока шпорна борозна (лат. sulcus calcarinus), передній - тім'яно-потилична борозна.
Борозни та звивини нижньої поверхні
На нижній поверхні лобової частини розташовується нюхова борозна (лат. sulcus olfactorius). Усередині від неї, між нею і нижньомедіальним краєм півкулі, лежить пряма звивина (лат. gyrus rectus). Її задній відділ доходить до передньої продірявленої речовини (лат. substantia perforata anterior). Назовні від борозни розташовується решта нижньої поверхні лобової частки, порізана короткими борошнями очних (лат. sulci orbitales), на ряд невеликих очних звивин (лат. gyri orbitales) .
Нижня поверхня скроневої частки глибокої борозна гіпокампа (лат. sulcus hippocampi) Відокремлена від ніжок мозку. У глибині борозни залягає вузька зубчаста звивина (лат. gyrus dentatus). Передній її кінець переходить у гачок, а задній - у стрічкову звивину (лат. gyrus fasciolaris) що залягає під валиком мозолистого тіла. Латерально від борозни знаходиться парагиппокампальна звивина (лат. gyrus parahippocampalis). Попереду ця звивина має потовщення як гачка (лат. uncus), а кзади продовжується в язичну звивину (лат. gyrus lingualis). Парагіппокампальну та язичну звивини з латерального боку обмежує колатеральна борозна (лат. sulcus collateralis), що переходить спереду в носову борозну (лат. sulcus rhinalis). Решту нижньої поверхні скроневої частки займають медіальна та латеральна потилично-скроневі звивини (лат. gyri occipitotemporales medialis et lateralis ), розділені потилично-скроневою борозна (лат. sulcus occipitotemporalis). Латеральна потилично-скронева звивинанижньолатеральним краєм півкулі відокремлюється від нижньої скроневої звивини.
Гістологія
Будова
Цитоархітектоніка (розташування клітин)
- молекулярний шар
- зовнішній зернистий шар
- шар пірамідальних нейронів
- внутрішній зернистий шар
- гангліонарний шар (внутрішній пірамідний шар; клітини Беца)
- шар поліморфних клітин
Мієлоархітектоніка (розташування волокон)
Кора півкуль головного мозку представлена шаром сірої речовини завтовшки в середньому близько 3 мм (1,3 – 4,5 мм). Найбільш сильно розвинена вона у передній центральній звивині. Велика кількість борозен і звивин значно збільшує площу сірої речовини головного мозку. У корі міститься близько 10-14 млрд. нервових клітин. Різні її ділянки, що відрізняються один від одного деякими особливостями розташування та будови клітин (цитоархітектоніка), розташування волокон (мієлоархітектоніка) та функціональним значенням, називаються полями. Вони являють собою місця вищого аналізу та синтезу нервових імпульсів. Різко окреслені межі між ними відсутні. Для кори характерне розташування клітин і волокон шарами.
Цитоархітектоніка
Пірамідні клітини різних шарів кори відрізняються розмірами та мають різне функціональне значення. Дрібні клітини є вставними нейронами, аксони яких пов'язують окремі ділянки кори однієї півкулі (асоціативні нейрони) або двох півкуль (комісуральні нейрони). Ці клітини зустрічаються у різних кількостях у всіх шарах кори. Особливо багата на них кора головного мозку людини. Аксони великих пірамідних нейронів беруть участь в утворенні пірамідних шляхів, що проектують імпульси у відповідні центри мозкового ствола і спинного мозку.
Нейрони кори розташовані нерізко відокремленими шарами. Кожен шар характеризується переважанням одного виду клітин. У руховій зоні кори розрізняють 6 основних шарів:
Кора півкуль головного мозку також містить потужний нейрогліальний апарат, що виконує трофічну, захисну, опорну та розмежувальну функції.
На медіальній та нижній поверхні півкуль збереглися ділянки старої, стародавньої кори, які мають двошарову та тришарову будову.
Молекулярний шар
Молекулярний шар кори містить невелику кількість дрібних асоціативних клітин веретеноподібної форми. Їхні аксони проходять паралельно поверхні мозку у складі тангенціального сплетення нервових волокон молекулярного шару. Основна маса волокон цього сплетення представлена розгалуженнями дендритів нейронів нижчих шарів.
Зовнішній зернистий шар
Зовнішній зернистий шар утворений дрібними нейронами діаметром близько 10 мкм, що мають округлу, незграбну і пірамідальну форму, і зірчастими нейронами. Дендрити цих клітин піднімаються у молекулярний шар. Аксони або йдуть у білу речовину, або, утворюючи дуги, також надходять у тангенційне сплетення волокон молекулярного шару.
Шар пірамідальних нейронів
Є найширшим у порівнянні з іншими шарами кори головного мозку. Він особливо добре розвинений у прецентральній звивині. Величина пірамідних клітин послідовно збільшується в межах 10-40 мкм від зовнішньої зони цього шару до внутрішньої. Від верхівки пірамідної клітини відходить головний дендрит, що розташовується у молекулярному шарі. Дендрити, що беруть початок від бічних поверхонь піраміди та її основи, мають незначну довжину та утворюють синапси із суміжними клітинами цього шару. Аксон пірамідної клітини завжди відходить від її основи. У дрібних клітинах він залишається у межах кори; аксон ж, що належить великій піраміді, зазвичай формує мієлінове асоціативне або комісуральне волокно, що йде в білу речовину.
Внутрішній зернистий шар
У деяких полях кори розвинений дуже сильно (наприклад, в зоні зорової кори). Однак в інших ділянках він може бути відсутнім (у прецентральній звивині). Цей шар утворений дрібними зірчастими нейронами. До його складу входить велика кількість горизонтальних волокон.
Гангліонарний шар (Внутрішній пірамідний шар; Клітини Беца)
Утворений великими пірамідними клітинами, причому область прецентральної звивини містить гігантські клітини, описані вперше російським анатомом В. А. Бецем в 1874 року (клітини Беца). Вони досягають у висоту 120 і завширшки 80 мкм. На відміну з інших пірамідних клітин кори гігантські клітини Беца характеризуються наявністю великих глибок хроматофільного речовини. Їхні аксони утворюють головну частину кортико-спінальних і кортико-нуклеарних шляхів і закінчуються на мотонейронах мозкового ствола і спинного мозку.
Мієлоархітектоніка
Серед нервових волокон кори півкуль головного мозку можна виділити:
Крім тангенціального сплетення молекулярного шару, на рівні внутрішнього зернистого та гангліонарного шарів розташовані два тангенціальні шари мієлінових нервових волокон і колатералей аксонів клітин кори. Вступаючи в синаптичні зв'язки з нейронами кори, горизонтальні волокна забезпечують широке поширення у ній нервового імпульсу.
Модуль
I, II, III, IV, V, VI - шари кори
Аферентні волокна
1. кортико-кортикальне волокно
2. таламо-кортикальне волокно
2а. зона поширення специфічних таламо-кортикальних волокон
3. пірамідні нейрони
3а. загальмовані пірамідні нейрони
4. гальмівні нейрони та їх синапси
4а. клітини з аксональним пензликом
4б. малі кошикові клітини
4 в. великі кошикові клітини
4г. аксоаксональні нейрони
4д. клітини з подвійним букетом дендритів (гальмівні нейрони, що гальмують)
5. шипикові зірчасті клітини, що збуджують пірамідні нейрони безпосередньо та шляхом стимуляції клітин з подвійним букетом дендритів
Досліджуючи кору великих півкульголовного мозку Я. Сентаготаї та представники його школи встановили, що її структурно-функціональною одиницею є модуль- Вертикальна колонка діаметром близько 300 мкм. Модуль організований навколо кортико-кортикального волокна, що є аксоном пірамідної клітини III шару (шару пірамідальних клітин) тієї ж півкулі (асоціативне волокно), або від пірамідальних клітин протилежного (комісуральне). У модуль входять два таламо-кортикальні волокна - специфічні аферентні волокна, що закінчуються в IV шарі кори на шипикових зірчастих нейронах і відходять від основи (базальних) дендритах пірамідальних нейронів. Кожен модуль, на думку Сентаготаї, поділяється на два мікромодулі діаметром менше 100 мкм. Загалом у неокортексі людини приблизно 3 млн модулів. Аксони пірамідальних нейронів модуля проектуються на три модулі тієї ж сторони і через мозолисте тіло за допомогою комісуральних волокон на два модулі протилежної півкулі. На відміну від специфічних аферентних волокон, що закінчуються в IV шарі кори, кортико-кортикальні волокна утворюють закінчення у всіх шарах кори, і, досягаючи I шару, дають горизонтальні гілки, що виходять далеко за межі модуля.
Крім специфічних (таламо-кортикальних) аферентних волокон, на вихідні пірамідальні нейрони збудливий вплив мають шипикові зірчасті нейрони. Розрізняють два типи шипикових клітин:
Гальмівна система модуля представлена такими типами нейронів:
Система пригнічення гальмівних нейронів:
Потужний збуджуючий ефект фокальних шипикових зірчастих клітин пояснюється тим, що вони одночасно збуджують пірамідні нейрони та клітину з подвійним букетом дендритів. Таким чином, перші три гальмівні нейрони гальмують пірамідні клітини, а клітини з подвійним букетом дендритів збуджують їх, пригнічуючи гальмівні нейрони.
Однак, також існують критичні та альтернативні концепції , що ставлять під сумнів модульну організацію кори великих півкуль та мозочка. Безумовно, впливом геть ці думки справило прогноз у 1985 р. й надалі відкриття 1992 р. дифузного об'ємного нейротрансмиттинга .
Резюме
Міжнейрональні взаємозв'язки нейронів кори великих півкуль головного мозку можна уявити так: вхідна (аферентна) інформація надходить з таламуса по таламо-кортикальних волокнах, які закінчуються на клітинах IV (внутрішнього зернистого) шару. Його зірчасті нейрони мають збуджуючий вплив на пірамідні клітини III (пірамідальних нейронів) і V (гангліонарних) шарів, а також на клітини з подвійним букетом дендритів, які блокують гальмівні нейрони. Клітини III шару утворюють волокна (асоціативні та комісуральні), які пов'язують між собою різні відділи кори. Клітини V та VI (мультиморфних клітин) шарів формують проекційні волокна, які йдуть у білу речовину та несуть інформацію іншим відділам центральної нервової системи. У всіх шарах кори знаходяться гальмівні нейрони, які відіграють роль фільтра шляхом блокування пірамідних нейронів.
Кора різних відділів характеризується переважним розвитком тих чи інших її верств. Так, у рухових центрах кори, наприклад, у передній центральній звивині, сильно розвинені III, V і VI і погано виражені II і IV шари. Це так званий агранулярний тип кори. З цих областей беруть початок низхідні провідні шляхи центральної нервової системи. У чутливих кіркових центрах, де закінчуються аферентні провідники, що йдуть від органів нюху, слуху та зору, слабо розвинені шари, що містять великі та середні пірамідні клітини, тоді як зернисті шари (II та IV) досягають свого максимального розвитку. Це гранулярний тип кори .
Цитоархітектонічні поля БродманаІ. Н. Філіпповим і С. А. Саркісовим були створені карти кори головного мозку, що включають 47 цитоархітектонічних полів.
Складовими частинами мозкових півкуль є мозковий плащ та підкіркові ганглії. Оточують їх бокові шлуночки.
Між правою півкулею та лівою проходить глибока поздовжня борозна. У її глибині розташовується мозолисте тіло. Його утворюють нервові волокна.
Кора великих півкуль головного представлена мозковим плащем. Ця сіра речовина сформована нервовими клітинами з відростками, що відходять від них, і вважається, що останні виконують для нейронів опорну функцію та беруть участь в обміні їх речовин.
Кора великих півкуль є найвищим, з філогенетичного погляду, наймолодшим формуванням ЦНС. Товщина її шару від півтора до трьох міліметрів. Кора великих півкуль має близько дванадцяти-вісімнадцяти мільярдів нейронів.
Її загальна поверхня зростає за рахунок наявності численних борозен. Вони поділяють поверхні півкуль на частки і опуклі звивини. У кожній півкулі знаходиться чотири частки. Вони формуються за рахунок трьох борозен: бічної, тім'яно-потиличної та центральної. В результаті утворюються потилична, скронева, тім'яна та лобна частки.
Остання розташована спереду від центральної борозни. Тіменна частка обмежується центральною борозеною попереду, внизу – бічною, тім'яно-потиличною – ззаду. Скроневу часткуобмежує глибока бічна борозна вгорі. Потилична частка розташована ззаду від тім'яно-потиличної.
Вище мозолистого тіла знаходиться До його складу входять проекційні, комісуральні та асоціативні волокна. Кора великих півкуль має двосторонній зв'язок з відділами ЦНС, що лежать нижче, за допомогою висхідних і низхідних шляхів. До їх складу входять проекційні волокна, що виходять за межі півкуль.
Окремі кіркові області мають функціональне різне значення. Разом з цим кора великих півкуль працює як єдине ціле. Однак у ній немає суворої функціональної локалізації. Досліди над тваринами показали, що після руйнування окремих ділянок у корі, після деякого періоду часу сусідні області починали виконувати функції зруйнованих ділянок. Цю особливість пов'язують із високою пластичністю клітин.
Кора півкуль приймає від рецепторних утворень доцентрові імпульси. Для кожного апарату рецепторів у ній відповідає ділянка, названа І.П. Павловим "кіркове ядро аналізатора". Області кори, в яких вони розташовуються, називаються сенсорними ділянками.
У задньоцентральній та передньоцентральній зонах кори розташовується ядерна область рухового аналізатора. В нього проводиться від рецепторів сухожиль, скелетних м'язіві суглобів збудження.
Область розташована позаду центральної борозни (в задньоцентральній зоні). Він пов'язаний з тактильною, больовою та температурною чутливістю.
Найбільшу площу займає область аналізаторів особи, голосового апарату, кистей рук. Найменша площавідведено представництву аналізаторів гомілки, стегна та тулуба.
У потиличній зоні розташована ядерна область у скроневій – слуховій. Область знаходиться поблизу бічної борозни.
Рух виникає внаслідок подразнення, що формується від взаємодії із сенсорними областями моторної зони кори. Розташована вона від центральної борозни допереду.
Ядерні області аналізаторів представлені у корі ділянками, у яких закінчується більшість їх провідних шляхів. За межами розташовуються розсіяні елементи. Вони здійснюється надходження імпульсів тієї ж рецепторів, що у ядро аналізатора.
Кора великих півкуль
Регенерація.
Сіра речовина дуже погано регенерує. Біла речовина здатна регенерувати, але цей процес дуже тривалий. Якщо збережено тіло нервової клітини. То волокна регенерують.
У ній здійснюється вищий функціональний аналіз подразників і синтез-то є прийняття осмислених рішень для усвідомленої рухової реакції. У КДМ розташовуються центральні (кіркові) відділи аналізаторів; проводиться остаточне диференціювання подразнення. Основна функція КДМ – мислення.
Розвивається із переднього мозкового міхура. У його стінці проліферують вентрикулярні клітини, з яких диференціюються гліобласти та нейробласти (перші 2 тижні). Поступово проліферація нейробластів знижується. З гліобластів утворюється радіальна глія, відростки клітин якої пронизують всю стінку нервової трубки. Нейробласти мігрують протягом цих відростків, поступово диференціюються в нейрони (16-20 тиждень). Спочатку закладаються крайні шари кори, та був між ними утворюються проміжні шари. Розвиток кори продовжується після народження і завершується до 16-18 років. У процесі розвитку утворюється велика кількість нервових клітин, особливо розвиваються міжнейронні синапси. Що веде до утворення рефлекторних дуг.
КГМ представлена платівкою сірої речовини завтовшки 3-5мм, яка зовні покриває великі півкулі. Вона містить ядра як полів. Чіткої межі між полями немає, вони переходять одна в одну. Сіра речовина відрізняється високим вмістом нервових клітин. До 17-20млрд. Вони всі мультиполярні, різного розміру, за формою переважають пірамідніі зірчасті нервові клітини. Особливості розподілу нервових клітин у мозку позначаються терміном архітектоніка. Для КДМ характерна пошарова організація, де класично виділяють 6 шарів, між якими немає чіткої межі. Зовні до КГМ лежить м'яка мозкова оболонка, яка містить піальні судини, які під прямим кутом впроваджуються в КГМ.
1. Молекулярний шар -порівняно широкий шар. Містить невелику кількість веретеноподібнихнейронів, що розташовуються горизонтально. Основний обсяг цього шару складають відростки (слабко мієлінізовані), які надходять з білої речовини, в основному з кори цього ж або інших ділянок кори мозку обох півкуль. Більшість розташовується горизонтально, вони утворюють велику кількість синапсів. Цей шар виконує асоціативнуфункцію цієї ділянки з іншими відділами цієї півкулі або іншої півкулі. У молекулярному шарі закінчуються збуджуючі волокна, що несуть інформацію від ретикулярної формації. Через даний шар збудлива неспецифічна імпульсація передається на нижчі шари.
2. Зовнішній зернистий шар порівняно вузький. Характеризується високою частотою розташування нервових клітин, переважають дрібні піраміднінейрони. Дендрити цих клітин йдуть у молекулярний шар, а аксони в КГМ цієї півкулі. Клітини забезпечують зв'язок з іншими ділянками кори цієї півкулі.
3. Пірамідний шар -Найширший шар. Містить піраміднінейрони-дрібні, середні (переважно), великі, які утворюють 3 підшари. Дендрити цих клітин досягають молекулярного шару, аксони частини клітин закінчуються в інших ділянках кори цієї півкулі або протилежної півкулі. Οʜᴎ утворюють асоціативні нервові шляхи. Виконують асоціативні функції. Частина нервових клітин-аксони великих пірамідних нейронів йдуть у білу речовину і беруть участь у освіті низхідних проекційних рухових шляхів. Цей шар виконує найпотужніші асоціативні функції.
4. Внутрішній зернистий шар - Вузький, містить дрібні зірчастіі піраміднінейрони. Їх дендрити досягають молекулярного шару, аксони закінчуються в корі мозку цієї півкулі або протилежної півкулі. При цьому частина відростків йде горизонтально в межах 4 шари. Виконує асоціативніфункції.
5. Гангліозний шар досить широкий, містить великі та середні піраміднінейрони. У ньому розташовуються гігантськінейрони (клітини Беца). Дендрити йдуть у шари, що виходять, досягають молекулярного шару. Аксони йдуть у білу речовину та утворюють низхідні рухові шляхи.
6. Поліморфний шар - Вуж, ніж гангліозний. Містить клітини різноманітні формою, але переважають веретеноподібнінейрони. Їх дендрити також йдуть у шари, що виходять, досягають молекулярного шару, а аксони вступають у білу речовину і беруть участь в освіті низхідних нервових рухових шляхів.
1-4 шари є асоціативними. 5-6 шари є проекційними.
До кори належить біла речовина. Воно містить нервові мієлінові волокна. Асоціативні волокна забезпечують зв'язок всередині однієї півкулі, комісуральні-між. різними півкулями, проекційні між відділами різного рівня.
У чутливих відділах кори (90%) містяться добре розвинені 2, 4 шари-зовнішній та внутрішній зернисті шари. Така кора відноситься до гранулярного типу кори.
У руховій корі добре розвинені проекційні шари, особливо 5. Це агранулярний тип кори.
Для КДМ характерна модульна організація. У корі виділяють вертикальні модулі, які займають усю товщину кори. У такому модулі в середній частині розташовується пірамідний нейрон, дендрит якого досягає молекулярного шару. Також є багато дрібних вставних нейронів, відростки яких закінчуються на пірамідному нейроні. Частина з них збуджують за функцією, а більша частина - гальмівні. У даний модуль з інших ділянок кори входить кортико-кортикальне волокно, що пронизує всю товщину кори, по ходу віддає відростки-колатералі на вставкові нейрони і невелика частина-на пірамідний нейрон і досягає молекулярного шару. Також в модуль входять 1-2 таламокортикальні волокна. Вони досягають 3-4 шару кори, розгалужуються і утворюють синапси з вставковими нейронами і пірамідним нейроном. Цими нервовими волокнами надходить аферентна збудлива інформація, яка через вставні нейрони, які регулюють проведення інформації, або безпосередньо надходить на пірамідний нейрон. Вона обробляється, утворюється ефекторний імпульс у початковому відділі аксона пірамідного нейрона, який відводиться від тіла клітини за аксоном. Цей аксон до складу нервового кортикоспінального волокна надходить в інший модуль. І так від модуля до модуля інформація передається з чутливих відділів рухову кору. Причому інформація йде як горизонтально, і вертикально.
КГМ відрізняється високою щільністю судинно-капілярної мережі та нервові клітини розташовуються в осередку з 3-5 капілярів. Нервові клітини високочутливі до гіпоксії. З віком відбувається погіршення кровопостачання та загибель частини нервових клітин та атрофія речовини мозку.
Нервові клітини кори мозку здатні регенерувати за збереження тіла нейрона. При цьому відновлюються пошкоджені відростки та утворюються синапси, за рахунок цього відновлюють нервові ланцюги та рефлекторні дуги.
Кора великих півкуль - поняття та види. Класифікація та особливості категорії "Кора великих півкуль" 2014, 2015.
Півкулі головного мозку Загальна будовапівкуль головного мозку. Півкулі дітей відрізняються від півкуль дорослих лише «дрібними деталями рельєфу». Приблизно до 9-10 років звивини та борозни півкуль займає таке саме положення, як і у дорослого. На поперечному та... .
"Мозок - це остання з таємниць природи, яка коли-небудь відкриється людині." англ. фізіолог Чарлз Скотт Шеррінгтон. «Асиметрія є основною властивістю життя.» Луї Пастер. Великі півкулі – парні утворення головного мозку. У людини вони досягають...
КОРА ПІВКУЛЬ ВЕЛИКОГО МОЗКУ
Кора півкуль великого мозкує вищий і найбільш складно організований нервовий центр екранного типу,діяльність якого забезпечує регуляцію різноманітних функцій організму та складні форми поведінки.
Кора утворена шаром сірої речовинитовщиною 3-5 мм на поверхні звивин (30%) та в глибині борозен (70%) загальною площею 1500-2500 см 2 при об'ємі близько 300 см 3 . Сіра речовина містить нервові клітини(близько 10-15 млрд.), нервові волокнаі клітини нейроглії(понад 100 млрд.).
На підставі відмінностей щільності розташування та будови клітин (цитоархітектоніки),ходу волокон (мієлоархітектоніки)і функціональних особливостейрізних ділянок кори в ній виділяють 52 нерізко розмежовані поля.
Нейрони кори - мультиполярні, різних розмірів і форм, включають понад 60 видів, серед яких виділено два основні типи - пірамідні та непірамідні.
Пірамідні клітиниспецифічний для кори півкуль тип нейронів; за різними оцінками, становлять 50-90% всіх нейроцитів кори. Від апікального полюса їхнього конусоподібного (на зрізах - трикутного) тіла, який звернений до поверхні кори, відходить довгий (апікальний) покритий шипиками дендрит,прямує в молекулярний шар кори, де він розгалужується (рис. 11-І). Від базальної та латеральних частин тіла вглиб кори та в сторони від тіла нейрона розходяться 5-16 коротших бічних (латеральних) дендритів,які, розгалужені, поширюються в межах того ж шару, де знаходиться тіло клітини. Від середини базальної поверхні тіла відходить довгий і тонкий аксон,що йде в білу речовину, яка на відстані 60-90 мкм починає давати колатералі. Розміри пірамідних нейронів варіюють від 10 до 140 мкм; розрізняють гігантські, великі, середні та малі пірамідні клітини.
Основна функція пірамідних клітин інтеграція всередині кори(середні та малі клітини) та утворення еферентних шляхів(гігантські та великі клітини).
Непірамідні клітинирозташовуються практично у всіх шарах кори, сприймаючи аферентні сигнали, що надходять, а їх аксони поширюються в межах самої кори, передаючи імпульси на пірамідні нейрони. Ці клітини дуже різноманітні та переважно є різновидами зірчастих клітин.Вони включають шипикі зірчасті, кошикові, аксо-аксонні клітини, клітини-"канделябри", клітини з подвійним букетом дендритів, горизонтальні клітини Кахаля, клітини Мартіноттіта ін. Основна функція непірамідних клітин - інтеграція нейронних ланцюгів усередині кори.
Цитоархітектоніка кори півкуль великого мозку. Нейрони кори розташовуються нерізко розмежованими шарами (пластинками),які позначаються римськими цифрами та нумеруються зовні всередину (див. рис. 11-11, А, Б).
/ - молекулярний шар розташовується під м'якою мозковою оболонкою; містить порівняно невелику кількість дрібних нейронів - горизонтальних клітин Кахаляз довгими розгалуженими дендритами, що відходять у горизонтальній площині від веретеноподібного тіла. Їхні аксони беруть участь в освіті тангенціального сплетення волоконцього шару. У молекулярному шарі є численні дендрити та аксони клітин більш глибоко розташованих шарів, що утворюють міжнейронні зв'язки.
II - Зовнішній зернистий шар утворений численними дрібними піраміднимиі зірчастими клітинами,дендрити яких розгалужуються і піднімаються в молекулярний шар, а аксони або йдуть у білу речовину, або утворюють дуги і також прямують у молекулярний шар.
III- піраміднийшар - значно варіює по ширині і максимально виражений у асоціативних та сенсомоторних областях кори.У ньому переважають пірамідні клітини,розміри яких збільшуються в глиб шару від дрібних до великих. Апікальні дендрити пірамідних клітин прямують у молекулярний шар, а латеральні утворюють синапси з клітинами даного шару. Аксони цих клітин закінчуються в межах сірої речовини або спрямовуються в білу. Крім пірамідних клітин, шар містить різноманітні Непірамідні нейрони.Шар виконує переважно асоціативні функції, зв'язуючи клітини як у межах цієї півкулі, так і з протилежною півкулею.
/V - внутрішній зернистийшар - широкий у зорової та слухової областях кори,а сенсомоторної області практично відсутня. Він освічений дрібними пірамідними та зірчастими клітинами.У цьому вся шарі закінчується переважна більшість таламічних аферентних волокон. Аксони клітин цього шару утворюють зв'язки з клітинами вище і нижче шарів кори.
V- гангліонарнішар освічений великими,а в області моторної кори (прецентральної звивини) - гігантськими пірамідними клітинами (Беца).Апікальні дендрити пірамідних клітин досягають I шару, утворюючи там верхівкові букети, латеральні дендрити поширюються в межах того ж шару. Аксони гігантських і великих пірамідних клітин проектуються на ядра головного та спинного мозку,найдовші з них у складі пірамідних шляхівдосягають каудальних сегментів спинного мозку. У V шарі зосереджено більшість кіркових проекційних еферентів.
VI - шар поліморфних клітин утворений різноманітними за формою нейронами (веретеноподібними, зірчастими, клітинамиMap-тінотті).Зовнішні ділянки шару містять більші клітини, внутрішні - дрібніші і рідко розташовані. Аксони цих клітин йдуть у білу речовину у складі еферентнихшляхів, а дендрити проникають до молекулярного шару. Аксони дрібних клітин Мартінотті піднімаються до кори і гілкуються в молекулярному шарі.
Мієлоархігектоніка кори півкуль великого мозку. Нервові волокна кори півкуль великого мозку включають три групи: 1) аферентні, 2) асоціативні та комісуральні та 3) еферентні волокна.
Кора великих півкуль є вищим відділом центральної нервової системи, який у процесі філогенетичного розвитку з'являється найпізніше і формується в ході індивідуального (онтогенетичного) розвитку пізніше за інші відділи мозку. Кора є шаром сірої речовини товщиною 2-3 мм, що містить в середньому близько 14 млрд. (від 10 до 18 млрд.) нервових клітин, нервові волокна і проміжну тканину (нейроглію). На поперечному її зрізі за розташуванням нейронів та їх зв'язків розрізняють 6 горизонтальних шарів. Завдяки численним звивинам та борознам площа поверхні кори досягає 0,2 м2. Безпосередньо під корою знаходиться біла речовина, що складається з нервових волокон, які передають збудження в кору та з неї, а також від одних ділянок кори іншим.
Коркові нейрони та їх зв'язки. Незважаючи на величезну кількість нейронів у корі, відомо дуже небагато їх різновидів.Основними типами їх є пірамідні та зірчасті нейрони.
...
В аферентної функції кори та в процесах перемикання збудження на сусідні нейрони основна роль належить зірчастим нейронам. Вони становлять у людини понад половину всіх клітин кори. Ці клітини мають короткі розгалужені аксони, що не виходять за межі сірої речовини кори, і короткі розгалужені дендрити. Зірчасті нейрони беруть участь у процесах сприйняття подразненні та об'єднанні діяльності різних пірамідних нейронів.
Пірамідні нейрони здійснюють еферентну функцію кори і внутрішньокіркові процеси взаємодії між віддаленими один від одного нейронами. Вони діляться на великі піраміди, від яких починаються проекційні, або еферентні шляхи до підкіркових утворень, і дрібні піраміди, що утворюють асоціативні шляхи до інших відділів кори. Найбільші пірамідні клітини - гігантські пірамідиБеца знаходяться в передній центральній звивині, в так званій моторній зоні кори. Характерна рисавеликих пірамід - їхня вертикальна орієнтація в товщі кори. Від тіла клітини вертикально вгору до поверхні кори спрямований найбільш товстий (верхівковий) дендрит, через який в клітину надходять різні аферентні впливи від інших нейронів, а вертикально вниз відходить еферентний відросток - аксон.
Численність контактів (наприклад, тільки на дендритах великої піраміди їх налічують від 2 до 5 тис.) забезпечує можливість широкого регулювання діяльності пірамідних клітин з боку безлічі інших нейронів. Це дозволяє координувати відповідні реакції кори (насамперед її моторну функцію) з різноманітними впливами із зовнішнього середовища та внутрішнього середовища організму.
Для кори великих півкуль характерна велика кількість міжнейронних зв'язків. З розвитком мозку людини після його народження збільшується кількість міжцентральних взаємозв'язків, особливо інтенсивно до 18 років.
...
Первинні, вторинні та третинні поля кори. Особливості будови та функціонального значення окремих ділянок кори дозволяють виділити окремі кіркові поля.
Розрізняють три основні групи полів у корі: первинні, вторинні та третинні поля.
Первинні поля пов'язані з органами почуттів та органами руху на периферії, вони раніше за інших дозрівають в онтогенезі, мають найбільші клітини. Це так звані ядерні зони аналізаторів, за І. П. Павловим (наприклад, поле больової, температурної, тактильної та м'язово-суглобової чутливості в задній центральній звивині кори, зорове поле в потиличній ділянці, слухове поле у скроневій ділянці та рухове поле в передній центральній звивині кори) (рис. 54). Ці поля здійснюють аналіз окремих подразнень, що надходять у кору від відповідних рецепторів. При руйнуванні первинних полів виникають так звана кіркова сліпота, кіркова глухота тощо. Поряд розташовані вторинні поля, або периферичні зони аналізаторів, які пов'язані з окремими органами тільки через первинні поля. Вони служать для узагальнення та подальшої обробки інформації, що надходить. Окремі відчуття синтезуються в них у комплекси, що зумовлюють процеси сприйняття. При поразці вторинних полів зберігається здатність бачити предмети, чути звуки, але їх не впізнає, не пам'ятає їх значення. Первинні та вторинні поля є і в людини, і тварин.
Найбільш далекі від безпосередніх зв'язків із периферією третинні поля, або зони перекриття аналізаторів. Ці поля є лише в людини. Вони займають майже половину території кори та мають великі зв'язки з іншими відділами кори та з неспецифічними системами мозку. У цих полях переважають найбільш дрібні та різноманітні клітини. Основним клітинним елементом тут є зірчасті нейрони. Третичні поля знаходяться у задній половині кори - на межах тім'яних, скроневих та потиличних її областей та у передній половині - у передніх частинах лобових областей. У цих зонах закінчується найбільше нервових волокон, що з'єднують ліве і права півкулітому роль їх особливо велика в організації узгодженої роботи обох півкуль. Третичні поля дозрівають в людини пізніше інших кіркових полів, вони здійснюють найскладніші функції кори. Тут відбуваються процеси вищого аналізу та синтезу. У третинних поляхна основі синтезу всіх аферентних роздратувань і з урахуванням слідів колишніх роздратування виробляються цілі та завдання поведінки. Відповідно до них відбувається програмування рухової діяльності. Розвиток третинних полів у людини пов'язують із функцією мови. Мислення (внутрішня мова) можливе тільки при спільної діяльностіаналізаторів, об'єднання інформації яких відбувається у третинних полях.
При вродженому недорозвиненні третинних полів людина неспроможна опанувати промовою (вимовляє лише безглузді звуки) і навіть найпростішими руховими навичками (не може одягатися, користуватися знаряддями праці тощо).
Сприймаючи та оцінюючи всі сигнали з внутрішнього та зовнішнього середовища, кора великих півкуль здійснює найвищу регуляцію всіх рухових та емоційно-вегетативних реакцій.
Функції кори великих півкуль. Кора великих півкуль виконує найскладніші функції організації пристосувальної поведінки організму у зовнішньому середовищі. Це насамперед функція вищого аналізу та синтезу всіх аферентних подразнень.
Аферентні сигнали надходять у кору різними каналами, у різні ядерні зони аналізаторів (первинні поля), та був синтезуються у вторинних і третинних полях, завдяки діяльності яких створюється цілісне сприйняття зовнішнього світу. Цей синтез лежить в основі складних психічних процесів сприйняття, уявлення, мислення. Кора великих півкуль є орган, тісно пов'язаний із виникненням у людини свідомості та регуляцією її суспільної поведінки. Важливою стороною діяльності кори великих півкуль є замикальна функція – утворення нових рефлексів та їх систем ( умовні рефлекси, Динамічні стереотипи-див. розділ XV).
Завдяки надзвичайно велику тривалість збереження в корі слідів колишніх роздратування (пам'яті) у ній накопичується величезний обсяг інформації. Це має велике значеннядля збереження індивідуального досвіду, який використовується за необхідності.
...
Експериментально показано, що у вищих представників тваринного світу після повного оперативного видалення кори вища нервова діяльністьрізко погіршується. Вони втрачають здатність тонко пристосовуватися до зовнішнього середовища та самостійно існувати в ньому.